

The Big Abstraction for AI
The Big Abstraction for AI
An Ecological Development Meta-Framework

Previously, I talked about “Nature-Inspired Development as an AI Abstraction.” In other words, a meta-framework of sorts for bio-inspired Artificial Intelligence—including cognitive architectures and Strong AI (sometimes overlapping with the definition of AGI).
“Strong” AI seeks to create artificial persons: machines that have all the mental powers we have, including phenomenal consciousness. “Weak” AI, on the other hand, seeks to build information-processing machines that appear to have the full mental repertoire of human persons1
Specifying the Unspecific
The concept of abstraction helps in levels of detail, so you can zoom in on one area but zoom out as needed without losing track of other contexts. Abstractions also make it more clear what exactly you’re mimicking from nature and to what degree.

Arguably all AI work inspired by nature involves abstracting a lot—but not all researchers / projects are up front about it or even think about that aspect. For instance your artificial neural net may have some vague high level similarities to a biological brain circuit, but it’s only able to model certain things and the other details are just hidden away and you don’t worry about it. The “neurons” themselves are nothing like biological neurons. The analog wetware of animals is abstracted and replaced with digital computing and so forth…
From a usability standpoint, it’s necessary to abstract at least some details if not the majority. Our conceptions of tech typically end up in layers, nested layers, parallel units and orthogonal units each with their own sets of layers. If I want to take a video with my phone I can’t be bothered at that moment with thinking about the code behind that program, how was it compiled to bytecode or machine code, how does it interact with the OS, how do the camera electronics work, how does the OS run on the CPU, the multiple cores in the CPU, how does that interact with the GPU (which might be in the same chip), how does the circuitry work within each core and so on. It’s not quite turtles all the way down but it feels like it.
As with usability, I think AI research requires abstractions but also needs to recognize and admit their existence.
And abstraction space exploration can not only shape our framework but also our meta-framework—how are we thinking about the topic itself, what theoretical framework are we operating in and how does that lead to design, implementation and experimentation?
What follows is based on a paper I wrote in 20132.
The Big Abstraction
Natural intelligence entangles mental life, behavior and environment.
Besides the research desire to achieve biological kinds of intelligence in artificial systems, a complimentary worthwhile goal is to expand the domains of AI to include more real world environments where humans and other animals normally operate. So, one might ask what are the causes of intelligence (and lapses thereof) in natural contexts?
A particular animal’s behavior at any given time can be explained at several simultaneous levels, which come in three kinds as categorized by Konner3:
Remote or evolutionary causation, which includes the genome which in turn is the result of phylogenetic constraints and ecological causes.
Developmental causation, which includes the embryonic period where the genome kickstarts ontogeny, formative early-environment effects and non-formative lifetime environmental effects.
Proximate or functional causation, which includes hormonal and metabolic effects, short-term physiology and elicitors. Short-term physiology, which refers primarily to neural circuits, is the immediate internal cause of behavior. Elicitors, or releasers, are stimuli which cause external behavior.
I propose to use abstractions of those causes as part of an ecological development approach.
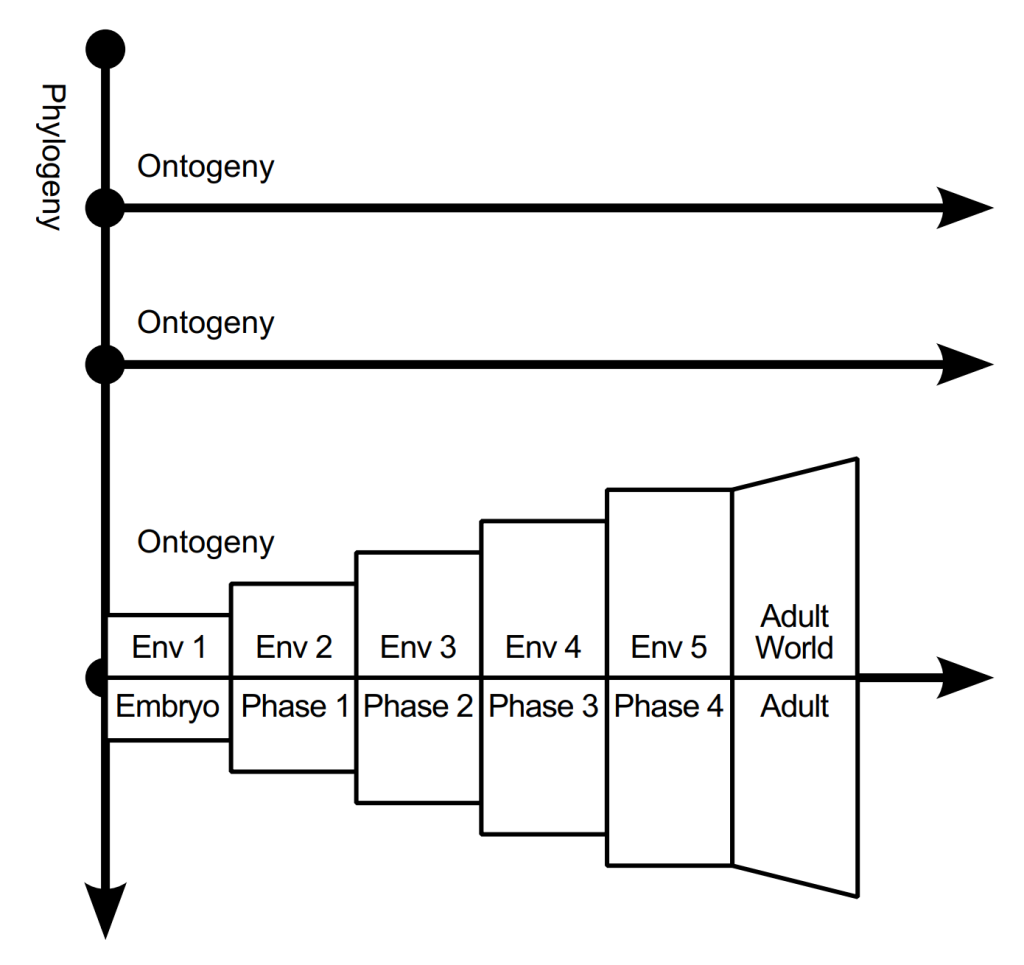
Development can be thought of as design in two dimensions: phylogeny and ontogeny. Phylogeny is evolutionary history, indicated by the vertical axis in the diagram below. An ontogeny occupies the lifespan of an organism; multiple ontogenies are represented in the diagram as horizontal axes. A special phase in ontogeny is embryogeny, where the genome instantiates the organism.
In reference to the aforementioned three kinds of causation, the remote/evolutionary kind of causation happens in phylogeny. Developmental causation and functional/proximate causation happen during ontogeny and embryogeny.
Biologically inspired development as a representation for a system has the potential benefits of compactness, scalability, self-organization, robustness in balance with adaptability, evolvability, fault tolerance and self-repair4. Whereas Floreano stated that development abstractions must include cellular mechanisms, I propose to abstract away those details.
Those aspects of biological development that are purely in support of the substrate will be abstracted out, such as myelination. However, it is quite possible for artificial substrate constraints to arise. But those should not be so great to make the endeavor impossible. It is certainly possible to evolve and grow software (I will discuss software development later). The embryonic transformation of information from phylogenetic space to ontogenetic space is actually easier in software than in biology since constraints can be entirely arbitrary in software. Even when using physical robots for embodiment, the entire process can be informational, e.g. autogenerating various electromechanical designs which are input to various machines in the world of the researcher to manufacture the robots which are injected back into the robot’s subset of the real world.
The essence of cellular mechanisms that remains in this abstraction is the modification of information systems without breaking existing functionality.
The development abstraction enables valuable AI research in two ways.
First, the aforementioned essence can be used as a requirement—for both phylogeny and ontogeny—to build intelligence as extensions to already working intelligence. So it is a way to build potentially complex AI with iterative simple changes.
Second, the abstraction recognizes the holistic nature of human-level intelligence. The intention is to inherently support ecological5,6 and embodied7 approaches, as well as overlapping enactive approaches, without necessarily excluding extended mind8 ideas that contrast with enactive ideas 9.
The enactive approach views perceptual experience as an activity of environmental exploration using sensorimotor dependencies and knowledge10. This approach would be difficult to experiment with using most AI abstractions and systems out of the box; only certain types of ALife and mobile autonomous robotics have even the minimal requirement of situated autonomous creatures. Development in an abstraction layer involving issues of an organism growing and living in an ecology provides the minimal configuration to observe and/or purposely design enactive perception and other forms of ecological cognition.
Agents in this development abstraction should typically be autopoietic systems, in which the organism is a phenomenon in itself as opposed to merely a role in population statistics, a notion which historically helped form the enactive approach11. An artificial organism developed as an autopoietic system ensures that the concept of organism is not just a label for a sum of parts12. Autopoietic artificial organisms are still developed and interdependent on their ecology, but they are also self-maintaining unified organizations.
Ontogenetic Frameworks
Ontogenetic space contains many kinds of things, such as the artificial organism (which may require an embryonic transformation mechanism to emerge from a digital space into a mixed digital-analog space), environmental agents and environmental artifacts (including amorphous things like “a mountain”). To work in this abstraction, one must have an ontogenetic framework which is composed of and attempts to keep track of these things (be they real or virtual).
There also must be meta-mechanisms for recording all aspects of the artificial organism’s development and interaction with the environment. The meta-mechanisms should store as much state information as possible at high frequency as well as additional substrate logging. Importantly, this information includes internal mental state and the actual mental architecture at a given sample time. Furthermore, these internal states can be correlated with external states. Meta-mechanisms can also be engineered for the ability to inject changes into the framework using new data or already recorded data. The ability to test a single variable change would be quite useful for a complex dynamical system.
Another aspect of ontogeny is the process of embryogeny, in which a genotype is transmogrified into a phenotype. A researcher could choose to use development during ontogeny but skip embryogeny by starting with a neonate. And the starting mental seed does not necessarily have to be evolved—it could come from any method. However, there may be benefits to using the two dimensions (phylogeny and ontogeny) fully in this abstraction. In one experiment, evolution with implicit (more like biology) artificial embryogeny outperformed not only other kinds of artificial embryogeny but also evolution with no embryogeny13.
Development and Learning Stages
Abstract development allows us to say what elements are plastic and at what time they are at a particular plasticity. In this light, learning and development are differentiated by the types of plasticity they use. An artificial organism learns with a plasticity that is more about content and new concepts built on older ones, whereas its development is more about building those underlying concepts and the very machinery needed to be able to learn.
Another ontogenetic framework tool that is particularly relevant is the inclusion of software/hardware created to be a causal contributor to an artificial organism’s development. A sequence of development phases could be established—such as designed microcosms of the adult ecological niche—to shape the plastic artificial mind. E.g., the first phase provides a very forgiving environment, the second one is a bit harder and has more challenging toys and so on until the organism reaches adulthood. Or, for learning, the phases could be established like the increasingly more difficult levels of a video game, presumably increasing the skill of the player at each level.
In the diagram shown earlier, the third ontogeny is expanded to indicate a four stage development pattern preceded by an embryogeny stage, culminating in an adult stage (“Env” means environment). The adult stage may include continual development and learning. In each stage, the environment has grown and the artificial organism has grown, including its internal cognitive architecture. Depending on the phenotype, its morphology may have changed at each stage, and if it did, that also affected its cognitive architecture.
I do not specify here what the design space is for an artificial organism to be able to develop and learn in increasingly more challenging stages. However, a good start is plasticity combined with curiosity combined with an environment of appropriate possible affordances. Autonomous development stages have been shown to emerge from a curious learning robot14.
Special Agents
The ecological development abstraction inherently supports agent interactions for development and/or learning. For instance, research into social intelligence could experiment with introductions of specifically designed interactive agents or homogeneous tribes of artificial organisms. Given that social complexity co-evolves with neocortex size in primates (and possibly other mammals)15, social experiments within ecologies could provide insight on mental structures and scaling for primate-level intelligence
Another special agent is the caregiver, typically providing maternal interactions in primates. It has been theorized that caregiver interactions are a primary causation of the human capacity for symbolic thought16. The artificial analogue would serve the purpose of triggering certain mental structures or competences such as emotionally-charged mental symbols for human-level conceptual cognition.
Agents, including but not limited to the aforementioned caregiver, could assist in training. It will become a matter of what static and dynamic elements of the environment to introduce at what times to achieve what developmental effects.
Iterative Evo-Devo
The four “secrets” of evolutionary innovation are17:
Tinkering (work with what is already present).
Multifunctionality.
Redundancy.
Modularity.
These essences should still be valid in a development abstraction, and they apply to both evolution and life history. Here I’m focusing on #1 (tinkering).
The practices of software engineering are relevant, and not just because AI is usually software. In software engineering the commonly used term development does not necessarily have anything to do with biological development. Yet, there are some similarities, namely with iterative methodologies.
Many software engineering projects use iterative development in which user stories or tasks are completed in small time boxes, sometimes called “sprints”. At some point, enough sprints have occurred to make the software product good enough for a release, after which development may or may not continue. The old Agile principles state that working software is the primary measure of success and that this working software should be delivered frequently18. One way to realize this is to make the software work at the end of every sprint, even if it is a very immature prototype. Although iterative is often associated with incremental, a purely incremental approach would not necessarily result in a working system after each increment. Besides being less brittle to change, iterative development ideally eliminates the need for later integration as all changes are also inherently integrations.
Nature uses iterative methods (aka tinkering) in phylogeny and ontogeny. Although ontogeny does not recapitulate phylogeny, there is a shared requirement: the changes of development (and trial-and-error learning) are always potential risks. If the change leaves the organism in a state where it’s not viable in its ecological/social context, it could die. Analogically, phylogeny risks fatal nonfunctional phenotypes when it mutates genes3.
Biological phylogenetic change is comparable to software development of artificial organism designs. Biological ontogenetic change is a similar phenomenon to the runtime changes of an artificial organism. Neither of these comparisons should be controversial; but I am saying that beyond those likenesses lie the ability to use the exact same methodology—accretion, which is a kind of iterative development.
In practice, software projects using iterative development methodologies can have varying requirements and mixtures of other methodologies. Here, a hard requirement of functional continuity at all moments in time regardless of maturity must be used.
The accretion is not merely addition, however, as it can modify older structures. The modifications should generally not remove any necessary functionality. Of course, “necessary” depends on the context—what ecological niche will the organism occupy? If there is a shift in context, then it may be plausible to also have a shift in functionalities. Removal in general can be a good method for refinement, e.g. biological removal of overly-connected neural pathways to leave the useful networks. Development of computer programs by humans have similar patterns where a program reaches a certain amount of bloat at which point refactoring, which may involve simplification and reduction, may be necessary to enable further development while maintaining the usefulness of the program.
Modification of ontogenetic structures can result in homologies; i.e. those structures which developed from the same sources but are slightly different in form or function. Homologies would be apparent when comparing an artificial organism to itself in the past (just as one can diff a revision of source code to a previous revision) or to an evolutionary relative. Homologies aren’t the only way to modify mind development; new structures have to be created as well.
There are some evolutionary biological tricks that could be used in artificial evolution of development as well, such as facultative adaptations (options that can be switched on/off by environmental stimuli) and heterochrony (timing of developments).
Another dynamic, which heterochrony can affect, is the altricial-precocial spectrum19. An example of a very precocial animal is a bird that hatches covered with down, eyes open, and can soon leave the nest. In contradistinction, the very altricial version would hatch with no down, eyes closed, and stay in the nest for a long time, dependent on its parents for food. An artificial organism could be made up of a mixture of both types—a particular mental structure (and behavior) could be precocial while another one is altricial. At the level of the entire organism in general, it could have a precocial embryogeny followed by altricial childhood development. The ecologies will determine what structures and skills need to be linked together on the altricial-precocial spectrum. Precocial mental mechanisms, such as bootstrapping learning, could be used in combination with—and be the cause of—altricial skills which learn reusable information chunks20.
Design of Design
For a development abstraction to make progress, it should not be reduced into just a narrow algorithm, such as evolving weights for artificial neural nets. The development abstraction uses life cycles in which autonomous mental architectures grow and learn. The mind is a system, but it is also an architecture, which at many levels of abstraction is composed of heterogeneous modules.
To make progress, Strong AI experimenters, if there are any left, should probably design ecological evo-devo frameworks which design minds. The heterogeneous modules and interfaces that tie them together should be a main focus of concern.
And it all should be in the context of iterative development of genotypes and phenotypes.
This is a shift from mindless approaches of machine learning (ML) and evolutionary ALife.
It was stated over two decades ago that behavioral robots and ALife had reached an impasse, and artificial creatures had never taken off like natural systems21.
This is not the mystery it may seem—it is because of a lack of meta-design in a development abstraction and a lack of architectural thinking.
But, one might argue, the whole point of most AI systems inspired by biology is to have the computer figure out the design for you, for instance by evolving the best fit-for-walking robots in micro-worlds.
So how does one reconcile this potential design paradox?
I suspect the answer is already known by many software developers: a design lives with the thing that is designed. Design is just a derivative view of an abstraction of something. And a document referred to as “the design” is not really the design—at best it’s a representation of a static concept of the design which may not even be feasible.
There is a balance (and a cycle) between upfront analysis/design versus coding implementations and testing them in the wild.
And there is a balance between coarseness of design at a given time—how detailed is it for a particular context?
A genotype must balance what details it codes for versus coding for structures that will figure out the details during ontogeny. Complex robust architectures that fit into their ecology emerge this way.
Taxonomies
A taxonomy of artificial embryogeny systems has been proposed before22 as well as taxonomies for robot skills23. It is conceivable that researchers could collectively contribute to a comparative taxonomy of evo-devo intelligence patterns that can be remixed into new working creatures and/or improved disembodied computer systems.
Comparisons and benchmarks can be difficult for AI, and it makes no sense to evaluate organisms from disparate niches with the same standardized test. Intelligence in one context does not necessarily result in intelligence in others.
Perhaps, instead, biological-style taxonomies and comparisons could be of use to AI practitioners using a developmental abstraction. Although the practitioners are also the designers at some meta-level, certainly they will observe unexpected emergent phenomena which could be compared and classified.
Taxonomies could define toolboxes of genotype modules which result in known good ontogenetic developments which in turn create known good mental structures or behaviors in particular ecological contexts. These modules could in turn can be used in new designs via composition.
Of course, modular AI doesn’t require an ecological developmental approach. But, for example in Duro et al24, the nature of idiosyncrasies between robots limits the re-usable modules to features of the least capable robot. And as with all modular systems, nontrivial re-use requires flexible interfaces. These two observations indicate that an adaptive developmental system might actually be more suited towards reuse of informational modules (although the “modules” may be genetic-like instructions on how to construct modules). As homologies, the reusable modules may have radically different forms depending on the artificial organism that uses it.
Conclusion
I have attempted to synthesize an AI abstraction based on phylogenetic and ontogenetic development. Many biological details such as cellular mechanisms are abstracted out. This approach involves design and observation of ecologies containing evolved artificial organisms. The intelligence of an artificial organism is not just inside its brain analogue, but also a property of an organism-environment system.
A useful ontogenetic framework might involve a series of increasingly more advanced environments (and possibly social interactions) to go along with an artificial organism’s child-like development and learning stages. Some of these stages may also include special agents, such as a caregiver to provide interactions needed for mental growth.
I also showed relevant relationships between iterative biological development and iterative software development. Designing artificial organisms will require a balance of architecture design and design of systems that automatically design other systems. Taxonomies could be made to enable comparisons of artificial evo-devo patterns amongst AI researchers.
Working with systems does not prevent focused research once the system is running. As a very speculative example, if vision is primarily an aspect of the sensorimotor behavior system, then the developmental approach enables an inherent methodology to always involve the sensorimotor-environment loop and avoid non-affordances. If a perception is not related to a bodily-mental use, then the organism will not develop that perception.
Enactive25, embodied26, and “more” Heideggerian27 approaches to intelligence and consciousness lead to the possibility that there cannot be humanlike cognition aside from an organism dynamically existing—in and as part of—its environmental niche. Therefore, an ecological development abstraction could be key to making humanlike artificial intelligence.
However, much meta-development and development remains to see if an ecological development abstraction is the right way to produce artificial creatures which live as natural creatures do, and if this is the right way to view intelligence.
Bringsjord, Selmer and Naveen Sundar Govindarajulu, “Artificial Intelligence”, The Stanford Encyclopedia of Philosophy (Fall 2022 Edition), Edward N. Zalta & Uri Nodelman (eds.), URL = <https://plato.stanford.edu/archives/fall2022/entries/artificial-intelligence/>.
Samuel H. Kenyon (2013). An Ecological Development Abstraction for Artificial Intelligence. Papers from the 2013 AAAI Fall Symposium.
Konner, M. 2010. The Evolution of Childhood: Relationships, Emotion, Mind. Cambridge, Mass.: Belknap Press of Harvard University Press.
Floreano, D., and Mattiussi, C. 2008. Bio-inspired Artificial
Intelligence: Theories, Methods, and Technologies. Cambridge, Mass.: MIT Press.Horton, T. E.; Chakraborty, A.; and St. Amant, R. 2012. Affordances for robots: a brief survey. Avant. The Journal of the Philosophical-Interdisciplinary Vanguard 3(2):70–84.
Montebelli, A.; Lowe, R.; and Ziemke, T. 2011. Energy constraints and behavioral complexity: The case of a robot with a living core. In AAAI Fall Symposium Series.
Anderson, M. L. 2003. Embodied cognition: A field guide. Artificial Intelligence 149(1):91 – 130.
Clark, A., and Chalmers, D. 1998. The extended mind. Analysis 58(1):10–23.
Gallagher, S., and Miyahara, K. 2012. Neo-pragmatism and enactive intentionality. In Schulkin, J., ed., Action, Perception and the Brain: Adaptation and Cephalic Expression. New York: Palgrave Macmillan.
Noë, A. 2004. Action in Perception. Cambridge, Mass.: MIT Press.
Froese, T. 2011. From second-order cybernetics to enactive cognitive science: Varela’s turn from epistemology to phenomenology. Systems Research and Behavioral Science 28(6):631–645.
Froese, T. 2011. Breathing new life into cognitive science. Avant. The Journal of the Philosophical-Interdisciplinary Vanguard 2(1):113–129.
Bentley, P., and Kumar, S. 1999. Three ways to grow designs: A comparison of evolved embryogenies for a design problem. In Genetic and Evolutionary Computation Conference, 35–43. Morgan Kaufmann.
Oudeyer, P.-Y.; Kaplan, F.; Hafner, V. V.; and Whyte, A. 2005. The playground experiment: Task-independent development of a curious robot. In Proceedings of the AAAI Spring Symposium on Developmental Robotics, 42–47.
Dunbar, R. I. M., and Shultz, S. 2007. Understanding primate brain evolution. Philosophical Transactions of the Royal Society B: Biological Sciences 362(1480):649–658.
Greenspan, S. I., and Shanker, S. 2004. The First Idea: How Symbols, Language, and Intelligence Evolved From Our Early Primate Ancestors to Modern Humans. Cambridge, MA: Da Capo Press.
Carroll, S. B. 2005. Endless Forms Most Beautiful: The New Science of Evo Devo. New York: Norton
Beck, K.; Beedle, M.; van Bennekum, A.; Cockburn, A.; Cunningham, W.; Fowler, M.; Grenning, J.; Highsmith, J.; Hunt, A.; Jeffries, R.; Kern, J.; Marick, B.; Martin, R. C.; Mellor, S.; Schwaber, K.; Sutherland, J.; and Thomas, D. 2001. Manifesto for agile software development. http://agilemanifesto.org.
Starck, J. M., and Ricklefs, R. E. 1998. Patterns of development: The altricial-precocial spectrum. In Starck, J. M., and Ricklefs, R. E., eds., Avian Growth and Development: Evolution Within the Altricial-Precocial Spectrum. New York, NY: Oxford University Press. 3–30.
Sloman, A., and Chappell, J. 2005. The altricial-precocial spectrum for robots. In Proceedings IJCAI ’05, 1187–1192.
Brooks, R. A. 2002. Flesh and Machines: How Robots Will Change Us. New York: Pantheon Books.
Stanley, K. O., and Miikkulainen, R. 2003. A taxonomy for artificial embryogeny. Artif. Life 9(2):93–130.
Huckaby, J., and Christensen, H. 2012. A taxonomic framework for task modeling and knowledge transfer in manufacturing robotics. In AAAI Workshops.
Duro, R. J.; Becerra, J. A.; and Santos, J. 2000. Improving reusability of behavior based robot cognitive architectures obtained through evolution. In Proceedings of the 6th ESA Workshop on Advanced Space Technologies for Robotics and Automation, 35–43. Noordwijk, The Netherlands: ESA.
Noë, A. 2009. Out of Our Heads: Why You Are Not Your Brain, and Other Lessons from the Biology of Consciousness. New York: Hill and Wang.
Vernon, D. 2008. Cognitive vision: The case for embodied perception. Image Vision Comput. 26(1):127–140.
Dreyfus, H. L. 2007. Why heideggerian ai failed and how fixing it would require making it more heideggerian. Philosophical Psychology 20(2):247–268.